 |
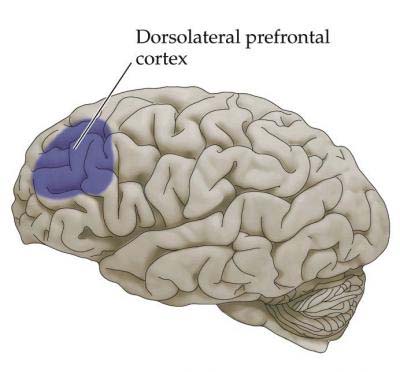
| Fig 1. From Kuo, Stokes, Murray & Nobre (2014) |
When you say ‘brain activity’, many people first think of
activity maps generated by functional magnetic resonance imaging (fMRI; see figure
1). As a non-invasive braining imaging method, fMRI has become the go-to workhorse
of cognitive neuroscience. Since the first papers were published in the early 1990s, there
has been an explosion of studies using this technique to study brain function, from basic perception to mind-reading for communicating with locked-inpatients or detecting lies in criminal investigations. At its best, fMRI
provides unparalleled access to detailed patterns of activity in the healthy human
brain; at its worst, fMRI could reduce to an expensive generator of 3-dimensional Rorschach images. To understand the relative strengths and weaknesses of fMRI, it is essential to understand exactly what fMRI measures. Without delving too deeply into
the nitty-gritty (see below for further reading), we will cover the basics that
are necessary for understanding the potential and limits of this ever popular and
powerful tool.
“fMRI does not directly measure brain activity”
First and foremost, electricity is the language of the
brain. At any moment, there are millions of tiny electrical impulses (action potentials) whizzing around your brain. At synaptic junctions, these impulses release
specific chemicals (i.e., neurotransmitters), which in turn modulate the electrical
activity in the next cell. This is the fundamental basis for neural communication.
Somehow, these processes underpin every thought/feeling/action you have ever
experienced. Our challenge is to understand how these electric events give rise
to these phenomena of mind.
However, fMRI does not exactly measure electrical activity (compare EEG, MEG, intracranial
neurophysiology); but rather it measures the indirect consequences of
neural activity (the
haemodynamic response). The pathway from neural activity to the fMRI activity
map is schematised in figure 2 below:
 |
| Fig 2. From Arthurs & Boniface (2002) |
 |
| Fig 3. From Oxford Sparks |
To summarise, let's consider three key principles: 1) neural activity is systematically
associated with changes in the relative concentration of oxygen in local blood
supply (figure 3); 2) oxygenated blood has different magnetic susceptibility relative to deoxygenated
blood; 3) changes in the ratio of oxygenated/de-oxygenated blood (haemodynamicresponse function; figure 4) can be inferred with fMRI by measuring the blood-oxygen-leveldependent (BOLD) response.
 |
| Fig 4. Haemodynamic response function |
So fMRI only provides an indirect measure of brain activity.
This is not necessarily a bad thing. Your classic thermometer does not directly
measure ‘temperature’, but rather the volume of mercury in a glass tube. Because
these two parameters are tightly coupled, a well calibrated thermometer does a nice
job of tracking temperature. The problem arises when the coupling is incomplete,
noisy or just very complex. For example, the haemodynamic response is probably
most tightly coupled to synaptic events rather than action potentials (see here). This
means certain types of activity will be effectively invisible to fMRI,
resulting in systematic biases (e.g., favouring input (and local processing) to
output neural activity). The extent to which coupling depends on unknown (or
unknowable) variability also limits the extent to which we can interpret the
BOLD signal. Basic neurophysiological research is therefore absolutely
essential for understanding exactly what we are measuring when we switch on the
big scanner. See here
for an authoritative review by Logothetis, a great pioneer in neural basis of
fMRI.
“spatial resolution”
Just like your digital camera, a brain scan can be defined
by units of spatial resolution. However, because the image is 3D, we call these
volumetric pixels, or voxels for short. In a typical scan, each voxel might cover
3mm3 of tissue, a volume that would encompass ~ 630,000 neurons
in cortex. However, the exact size of the voxel only defines the theoretically maximal
resolution. In practice, the effective resolution in fMRI also depends on the
spatial specificity of the hemodynamic response, as well as more practical considerations
such as the degree of head movement during scanning. These additional factors can add substantial spatial distortion or blurring. Despite these limits, there are few methods with superior spatial resolution. Intracranial recordings can measure activity with excellent spatial precision (even isolating activity from single cells), but this invasive procedure is limited to animal models or very specific clinical conditions that require this level of precision for diagnostic purposes (see here). Moreover, microscopic resolution isn't everything. If we focus in too closely without seeing the bigger picture, there is always the danger of not seeing the forest for all the trees. fMRI provides a good compromise between precision and coverage. Ultimately, we need to bridge different levels of analysis to capitalise on insights that can only be gained with microscopic precision and macroscopic measures that can track larger-scale network dynamics.
“snapshot is more like a long exposure photograph”
 |
| Fig 5. Wiki Commons |
Every student in psychology or neuroscience should be able to tell you that fMRI has good spatial
resolution (as above), but poor temporal resolution. This is because the haemodynamic
response imposes a fundamental limit on the time-precision of the measurement. Firstly, the peak response is delayed by approximately 4-6 seconds. However, this doesn’t really matter for offline analysis, because
we can simply adjust our recording to correct for this lag. The real problem is
that the response is extended over time. Temporal smoothing makes it difficult
to pinpoint the precise moment of activity. Therefore, the image actually
reflects an average over many seconds. Think of this like a very long long-exposure
photograph (see figure 5), rather than a snapshot of brain activity. This makes it very
difficult to study highly dynamic mental processes – fast neural
processes are simply blurred. Methods that measure electrical activity more directly have inherently higher temporal resolution (EEG, MEG, intracranial neurophysiology).
“too much data to make sense of”
A standard fMRI experiment generates many thousands of
measures in one scan. This is a major advantage of fMRI (mass
simultaneous recording), but raises a number of statistical challenges. Data
mining can be extremely powerful, however the intrepid data explorer will inevitably
encounter spurious effects, or false positives (entertain yourself with some fun
false positives here).
This is more of an embarrassment of riches, rather than a
limit. I don’t believe that you can ever have too much data, the important
thing is to know how to interpret it properly (see here). Moreover, the same problem applies to other data-rich measures of brain activity. The solution is not to limit our recordings, but to improve our analysis approaches to the multivariate problem that is the brain (e.g., see here).
“too many free parameters”
There are many ways to analyse an fMRI dataset, so which do
you choose? Especially when many of the available options make sense and can be easily
justified, but different choices generate slightly different results. This dilemma will be familiar to anyone who has ever analysed fMRI. A recent paper identified 6,912 slightly different paths
through the analysis pipeline, resulting in 34,560 different sets of results.
By fully exploiting this wiggle room, it should be possible to generate almost any
kind of result you would like (see here
for further consideration). Although this flexibility is not strictly a limit
in fMRI (and certainly not unique to fMRI), it is definitely something to keep in mind when interpreting what you read in the fMRI literature. It is important to define
the analysis pipeline independently of your research question, rather than try
them all and choose the one that gives you the ‘best’ result. Otherwise there
is a danger that you will only see what you want to see (i.e., circular analysis).
“…correlation, not causation”
It is often pointed out the fMRI can only provide correlational evidence. The same can be said for any other measurement technique. Simply because a certain
brain area lights up with a specific mental function, we cannot be sure that the observed activity actually caused the mental event (see here). Only an interference
approach can provide such causal evidence. For example, if we ‘knock-out’ a
specific area (e.g., natural
occurring brain damage, TMS, tDCS, animal ablation studies, optogenetics), and observe a specific impairment
in behaviour, then we can infer that the targeted area normally plays a causal role. Although this is strictly correct, this does not necessarily
imply the causal methods are better. Neural recordings can provide enormously rich
insights into how brain activity unfolds during normal behaviour. In contrast,
causal methods allow you to test how the system behaves without a specific area. Because
there is likely to be redundancy in the brain (multiple brain areas capable of performing
the same function), interference approaches are susceptible to missing
important contributions. Moreover, perturbing the neural system is likely to
have knock-on effects that are difficult to control for, thereby complicating
positive effects. These issues probably deserve a dedicated post in the future.
But the point for now is simply to note that one approach is not obviously
superior to the other. It depends on the nature of the question.
“…the spectre of reverse inference”
A final point worth raising is the spectre of reverse inference
making. In an influential review
paper, Russ Poldrak outlines the problem:
The usual kind of inference that is drawn from neuroimaging data is of the form ‘if cognitive process X is engaged, then brain area Z is active’. Perusal of the discussion sections of a few fMRI articles will quickly reveal, however, an epidemic of reasoning taking the following form:
- In the present study, when task comparison A was presented, brain area Z was active.
- In other studies, when cognitive process X was putatively engaged, then brain area Z was active.
- Thus, the activity of area Z in the present study demonstrates engagement of cognitive process X by task comparison A.
This is a ‘reverse inference’, in that it reasons backwards from the presence of brain activation to the engagement of a particular cognitive function.
Reverse inferences are not a valid from of deductive reasoning,
because there might be other cognitive functions that activate the brain area.
Nevertheless, the general form of reasoning can provide useful information,
especially when the function of the particular brain area is relatively specific and particularly well-understood. Using accumulated knowledge to interpret new findings is necessary for theory building. However, in the asbence of a strict one-to-one mapping between structure and function, reverse inference is best approached from a Bayesian
perspective rather than a logical inference.
Summary: fMRI is one of the most popular methods in cognitive
neuroscience, and certainly the most headline grabbing. fMRI provides unparalleled
access to the patterns of brain activity underlying human perception, memory
and action; but like any method, there are important limitations. To appreciate
these limits, it is important understand some of the basic principles of fMRI. We
also need to consider fMRI as part of a broader landscape of available techniques,
each with their unique strengths and weakness (figure 6). The question is not so much: is
fMRI useful? But rather: is fMRI the right tool for my particular question.
 |
| Fig 6. from Sejnowski, Churchland and Movshon, 2014, Nature Neuroscience |
Further reading:
Oxford Sparks (see below for video demo)
Key references
Arthurs, O. J.,
& Boniface, S. (2002). How well do we understand the neural origins of the
fMRI BOLD signal? Trends Neurosci, 25(1),
27-31. doi: Doi 10.1016/S0166-2236(00)01995-0
Logothetis, N. K. (2008). What we can do and what we cannot do with
fMRI. Nature, 453(7197), 869-878.
doi: DOI 10.1038/nature06976
Poldrack, R. A. (2006). Can cognitive processes be inferred from
neuroimaging data? Trends Cogn Sci, 10(2),
59-63. doi: DOI 10.1016/j.tics.2005.12.004
Sejnowski, T. J., Churchland, P. S., & Movshon, J. A. (2014).
Putting big data to good use in neuroscience. Nat Neurosci, 17(11), 1440-1441.
Fun demonstration from Oxford Sparks:





























{kind=link}
{kind=link}